Hits: 184


















































Fotografie, akvaristika, rodina a iné

Slovenská literatúra
Česká literatúra
Cudzojazyčná literatúra
Technika
Prírodné akvária
Ryby
Rastliny
Ekológia
Živorodky
Závojnatky
Kaprozúbky
Cichlidy
Diskusy
Tanganika
Malawi
Victoria
Afrika
Južná Amerika
Stredná Amerika
Spoločenstvo rýb, teda aj rastliny, mikroorganizmy, všetko živé v nádrži považujeme za biocenózu. Cenóza je spoločenstvo. Nemožno jednoznačne oddeliť jednotlivé časti, faktory, ktoré tvoria biocenózu. Biocenóza, spolu z neživými súčasťami nádrže tvoria ekosystém. Avšak možno hovoriť o ekosystéme akvária, ale aj o ekosystéme filtra, či kvapky vody. V akvaristike sa tieto pojmy veľmi nepoužívajú, iste aj preto lebo ide umelé ekosystémy, ktoré sú úzko závislé od energetických vstupov človeka. Spomínam ich, pretože sa s nimi napriek môžeme v akvaristike stretnúť. Napokon aj pri opise prírodných lokalít. Najmä v prírodných lokalitách je jasne vidieť vplyv biotických (živých) a abiotických (neživých) faktorov života rýb a rastlín. Len keď vezmem do úvahy geologické pomery – tie sú v akváriu zväčša absolútne popierané.
Z hľadiska prispôsobenia na kolísanie ekologických faktorov rozlišujeme druhy stenoekné a druhy euryekné. Stenoekné druhy znášajú malé kolísanie a euryekné druhy veľké kolísanie hodnôt. Známe pancierníčky Corydoras sa prispôsobili svojmu prostrediu natoľko, že dýchajú atmosférický vzduch črevnou sliznicou. Obdobne labyrinky dýchajú labyrintom atmosférický kyslík atď..
Podľa trofických parametrov sú rastliny producenti hmoty, živočíchy (teda aj ryby) sú konzumentmi. Mikroorganizmy spracovávajúce hmotu sú rozkladačmi – dekompozitormi. Podľa zdroja energie rozlišujeme organizmy na autotrofné – prijímajúce energiu za pomoci svetla a heterotrofné – spracúvajúce organickú, a neorganickú hmotu. Aj medzi rybami existujú rôzne vzťahy s ich okolím. Tento vzťah a ich usporiadanie skúma práve ekológia. Pre akvaristu je samozrejme najzaujímavejší vzťah ryba – ryba. Prípadne ryba – substrát dna – voda.
Technika výrazne v akváriu napomáha, až zabezpečuje život v akváriu. Treba si uvedomiť, že akvárium je umelý systém, ktorý je bez vstupov človeka len veľmi ťažko predstaviteľný. Medzi základné faktory ovplyvňujúce rast vodných rastlín patrí svetlo, dostupnosť živín, samotná voda, substrát, v prírode aj pôda. Vzťah existuje aj medzi rybami a rastlinami, vzájomne medzi na seba vplývajú. Rastliny dokážu tvoriť správnu mikroklímu pre ryby, poskytujú neraz možnosť úkrytov, no niekedy aj potravy. Faktor svetla rozdeľuje rastliny na tieňomilné a svetlomilné. Situácia je podobná ako v lese, kde zohráva svoju úlohu zápoj korún stromov, resp. kry v trópoch, epifity prepúšťajú na spodnú vrstvu nad hrabankou neraz iba 1% svetla. V prípade vodných rastlín, „zápoj“ tvoria rastliny na hladine, ktoré sú vyslovene svetlomilné. Svetlo pohlcujú aj plávajúce rastliny, vyššie rastliny a na koniec sa dostane aj na nízke rastliny dna, ktoré sú však tiež často svetlomilné. V prírode je síce primárnym zdrojom svetla slnko, ktorého svetlo je oveľa kvalitnejšie a intenzívnejšie, ale je pohlcované aj nad vodou často lesom, pobrežnou vegetáciou. A samozrejme nemožno zabudnúť na pohlcovanie svetla riasami, autotrofnými mikroorganizmami a samotnou vodou. Medzi tieňomilné rastliny patria: Anubias, Cryptocoryne, Vesicularia dubayana.
Za určitých okolností môže dôjsť v akváriu ku otrave. Zvyčajne ide o otravu amoniakom napr. spôsobenú vysokou hladinou organického odpadu, alebo o otravu nejakými kovmi z dekorácie. No zaujímavým spôsobom môže dôjsť k otrave aj vplyvom iného spracovania potravy tráviacim traktom inými druhmi rýb. Známe sú v tomto karasy, ktorých exkrementy sú pre iné druhy rýb jedovaté.
Konkurencia je známy termín. Konkurencia je hybnou silou vývoja. Jej prejavy sú pozorovateľné v nespočetnom množstve podôb aj u rýb a rastlín a ostatných organizmov v našich akváriách. Nemožno hovoriť v niektorých prípadoch o celkom normálnych prejavoch, pretože akvaristi zväčša iba napodobňujú prírodu. Vzorce platiace v prírode sú často pozmenené. Jeden z prípadom, kde v plnej miere obyčajne nemôžeme vidieť konkurenciu je potravná konkurencia. V akváriu si naši chovanci potravu vyhľadávajú len málo a na malom priestore. Preto nevznikajú také silné konkurenčné javy ako vo voľnej prírode. Konkurencia u rýb v akváriu sa prejavuje najmä pri zaujatí teritória a pri rozmnožovacích aktivitách. Konkurencia sa viac prejavuje u rastlín. Agresivitou sa medzi sebou vyznačujú samce bojovníc, ktoré zvádzajú medzi sebou neľútostné súboje. Najmä v prírode, keďže sa táto kombinácia v akváriu neodporúča. Na to, aby sme si to overili, môžeme použiť zrkadlo.
Vzťah koristi a dravca (predátora) je pozorovateľný aj v akváriu, niekedy to neznalého až šokuje. Keď cyklop, malý kôrovec dokáže vytvoriť na ryby tak silný predačný tlak, že mu plôdik rýb dokáže podľahnúť. Cyklop dokáže poter doslova uštípať. Typické dravce sú napr. šťučky loviace vodný hmyz, menšie ryby, niektoré cichlidy loviace ryby ako napr. juhoamerický Astronotus ocellatus, africké druhy rodu Nimbochromis. Tieto relatívne väčšie druhy rýb dorastajúce viac ako 20 cm, používajú zaujímavú techniku, kedy simulujú mŕtvolu nahnutú na dne. No ak sa do ich blízkosti priblíži menšia ryba, kaligono, ako nazýva tieto ryby domorodé obyvateľstvo okolia jazera Malawi, zrazu „ožijú“ a bleskovo sa snažia zmocniť svojej koristi. Ak zúžim tému na fakt, že korisť aj dravec sú ryby, podľa techniky lovu sa dajú rozlíšiť rôzne techniky lovu, ktoré ryby dodržujú. Niektoré ryby napádajú druhú odpredu, od hlavy, niektoré odzadu od chvostu, iné napádajú bok. Pravda, niektorým to je jedno. Dravá ryba je schopná viac-menej skonzumovať tak vysokú rybu, ako veľký je priemer jej oka. Samozrejme existujú výnimky.
Medzi suchozemskými rastlinami existuje jav známy ako alelopatia. Niektoré organizmy, resp. rastliny sa neznášajú do takej miery, že sú schopné sa likvidovať. Známy je tým orech, agát. Medzi vodnými rastlinami nebol vraj tento jav vedecky popísaný, osobne si myslím, že prirodzené vlastnosti vody priamo nahrávajú tomu, aby bol chemický boj medzi rastlinami intenzívnejší. Napr. známy český pestovateľ rastlín alelopatiu popisoval a neskôr tvrdil opak. Takmer v každej základnej akvaristickej literatúre sa možno dočítať, že do jedného akvária si zadovážte radšej zopár druhov rastlín, ako z každého dostupného druhu 1 – 2 jedince. Samozrejme to nie je len otázka boja medzi rastlinami, ale aj otázka vhodného substrátu pre ten ktorý druh, vhodného zloženia vody, použitej filtrácie, atď. V každom prípade biologické procesy jednotlivých rastlín a ekosystému akvária, prípadne vodných tokov, jazier či morí je studňa plná otázok (aj nevypovedaných samozrejme) a prekvapivých odpovedí. Nie je to vôbec také jednoduché, že by sme vzali nejakú rastlinu, zasadili a čakali že bude „rásť ako z vody“. Možnosti nádrže akvaristu sú obmedzené, napokon aj možnosti odbúravania látok v akvária sú priestorovo obmedzenejšie.
Voda poskytuje plynulejší prechod, väčšie rozptýlenie látok do priestoru, preto si myslím, že alelopatické prejavy sa musia prejaviť častejšie ako u rastlín na suchej zemi. Považujem to za ekologickú analógiu ku obranným mechanizmom, ku verbálnym prejavom nevôle, ku konkurenčným prejavom živočíchov. Je však možné, že substrát v nádrži neposkytuje toľko možností ako substrát v prírode a preto sa alelopatia ľahšie popíše práve v umelej nádrži. Pretože v akváriu skôr príde k prejavu náhleho stavu, najmä pre obmedzený priestor. Vo svojej praxi som sa stretol s prípadom, kedy som pestoval Cryptocoryne affinis v počte asi osem jedincov a pomerne veľký Echinodorus. Iné rastliny tam neboli. Pomerne uspokojivo rástli aspoň dva roky. Avšak raz, behom dvoch dní sa doslova všetky kryptokoryny rozpadli. Nezostalo z nich takmer nič, stonka sa oddelila od koreňa. Jediné čo z kryptokorýn zostalo, bol koreňový systém, z ktorého som následne kryptokoryny ďalej pestoval. Echinodorus rástol pokojne ďalej. Podobná situácia sa mi neskôr zopakovala znovu, v kombinácii s inou rastlinou. Neverím, žeby šlo o známu kryptokorynovú chorobu, pretože neviem o tom, že by sa iné podmienky sa menili. Šlo o prejav chemického boja, ktorý sa viedol zrejme najmä bohato rozvetvenými koreňmi oboch druhov rastlín.
Podľa prítomnosti kyslíka rozlišujeme dva základné procesy – anaeróbne a aeróbne. Anaeróbne procesy prebiehajú bez prístupu kyslíka, naopak aeróbne za prístupu kyslíka. S týmto popisom sa stretávame najmä pri rozklade hmoty. Aeróbne procesy aj anaeróbne na konci potravného reťazca zabezpečujú baktérie. Z trofického hľadiska rozlišujeme autotrofné baktérie, ktoré menia hmotu – zväčša anorganické látky za prístupu svetla a heterotrofné baktérie využívajú energiu zložitých organických zlúčením bez prístupu svetla tak. Pri anaeróbnom spracovaní dochádza aj ku štiepeniu na alkohol, metán, sulfán, na produkty jedovaté a teda nežiaduce pre život rastlín a rýb v akváriu. Preto je nutné dbať o dostatok neviazaného kyslíka v našom akváriu. Baktérie hmotu mineralizujú, táto energia je opäť transformovaná do vody, do koreňovej sústavy rastlín, kde sa opäť stáva prípadným začiatkom kolobehu látok.
The community of fish, including plants, microorganisms, and all living organisms in the tank, is considered a biocenosis. A cenosis is a community. It is not possible to clearly separate individual parts, factors that make up the biocenosis. The biocenosis, together with the inanimate components of the tank, forms an ecosystem. However, it is possible to talk about the aquarium ecosystem, as well as the filter or water droplets. In aquarium hobby, these terms are not widely used, perhaps because they represent artificial ecosystems that are closely dependent on human energy inputs. I mention them because we can encounter them in the context of describing natural habitats. Especially in natural habitats, the influence of biotic (living) and abiotic (non-living) factors on the life of fish and plants is clearly visible. Only when taking into account geological conditions – which are mostly completely negated in the aquarium.
From the perspective of adaptation to the fluctuation of ecological factors, we distinguish between stenoecious and euryecious species. Stenoecious species tolerate small fluctuations, while euryecious species tolerate large fluctuations in values. Well-known armored catfish Corydoras have adapted to their environment to the extent that they breathe atmospheric air through their intestinal mucosa. Similarly, labyrinth fish breathe atmospheric oxygen using their labyrinth organ, etc.
According to trophic parameters, plants are biomass producers, animals (including fish) are consumers. Microorganisms processing matter are decomposers. Based on the source of energy, organisms are classified as autotrophic – obtaining energy using light, and heterotrophic – processing organic and inorganic matter. Among fish, there are various relationships with their environment. The study of these relationships and their organization is the focus of ecology. For the aquarium hobbyist, the most interesting relationship is, of course, the interaction between fish or between fish, the substrate, and water.
Technology significantly contributes to the functioning of the aquarium and ensures life within it. It’s essential to realize that an aquarium is an artificial system that is challenging to conceive without human inputs. Among the fundamental factors influencing the growth of aquatic plants are light, nutrient availability, water itself, substrate, and, in nature, soil. There is also a relationship between fish and plants, and they mutually influence each other. Plants can create the right microclimate for fish, providing hiding places and sometimes even food.
The factor of light divides plants into shade-tolerant and light-loving. The situation is similar to a forest where the canopy of tree crowns plays a role, or in tropical areas, epiphytes allow only about 1% of light to reach the lower layer above the ground. In the case of aquatic plants, the “canopy” includes plants at the water surface, which are explicitly light-loving. Light is also absorbed by floating plants, taller plants, and eventually reaches the lower plants on the substrate, which are often also light-loving. Although sunlight is the primary source of light in nature, and its light is much higher in quality and intensity, it is often absorbed above the water by forests or coastal vegetation. And, of course, light is absorbed by algae, autotrophic microorganisms, and the water itself. Shade-tolerant plants include Anubias, Cryptocoryne, Vesicularia dubayana.
Under certain circumstances, aquariums can experience poisoning. It typically involves ammonia poisoning, for example, caused by a high level of organic waste, or poisoning by certain metals from decorations. Interestingly, poisoning can also occur due to the influence of the digestive tracts of other fish processing food differently. Carps are known for their excrements being toxic to other fish species in this regard.
Competition is a well-known term and a driving force in evolution. Its manifestations are observable in countless forms among fish, plants, and other organisms in our aquariums. In some cases, we can’t speak of entirely normal behaviors, as aquarium enthusiasts generally mimic nature, often with altered patterns. One case where we might not fully observe competition is in food competition. In the aquarium, our inhabitants seek food in a limited space. Consequently, strong competitive behaviors do not emerge as in the wild. Fish competition in the aquarium is mainly evident in territorial disputes and during reproductive activities. However, competition is more pronounced among plants. Males of betta fish, for example, display aggression by engaging in fierce battles. This is especially observed in nature, as such a combination is not recommended in aquariums. To confirm this, one can use a mirror.
The relationship between prey and predator is observable in the aquarium, sometimes shocking to the uninformed. For instance, a small copepod can create such a strong predatory pressure on fish that the fry of the fish can succumb. Copepods can literally nip at fish fry. Typical predators include pike hunting aquatic insects, smaller fish, and certain cichlids that prey on other fish, such as the South American Astronotus ocellatus or African species of the genus Nimbochromis. These relatively larger fish, growing over 20 cm, use an interesting technique of simulating a dead fish lying on the bottom. However, if a smaller fish, a “kaligono” as the native population around Lake Malawi calls these fish, approaches them, they suddenly “come to life” and swiftly try to seize their prey. Narrowing down the topic to the fact that both prey and predator are fish, various hunting techniques can be distinguished based on the fish’s behavior. Some attack from the front, from the head, some from behind, from the tail, and others attack from the side. Of course, some fish are indifferent to these distinctions. A predatory fish can consume a fish whose diameter is roughly the size of its eye. Naturally, there are exceptions.
Among terrestrial plants, there is a phenomenon known as allelopathy, where certain organisms or plants exhibit intolerance to each other to the extent of being capable of self-destruction. While this phenomenon has been scientifically described among land plants, it is believed that similar chemical warfare might occur more intensively among aquatic plants due to the natural properties of water. Some aquarists recommend having a variety of plant species in an aquarium rather than a few individuals of each species to avoid potential conflicts. However, this is not just a matter of plant competition but also involves considerations like suitable substrate, water composition, filtration methods, and more. The biological processes of individual plants and the aquarium ecosystem, as well as watercourses, lakes, or seas, are a wellspring of questions, some of which remain unanswered, and surprising answers.
Water provides a smoother transition and greater dispersion of substances into space. Therefore, allelopathic effects may occur more frequently among aquatic plants than their terrestrial counterparts. This can be viewed as an ecological analogy to defense mechanisms, verbal expressions of discontent, or competitive behaviors among animals. However, it is possible that the aquarium substrate doesn’t provide as many options as natural substrates, making allelopathy easier to observe in artificial tanks. In aquariums, sudden situations can arise, especially due to limited space.
The presence of oxygen distinguishes two fundamental processes — anaerobic and aerobic. Anaerobic processes occur without oxygen, while aerobic processes occur in the presence of oxygen. These descriptions are often encountered in the decomposition of matter. Both aerobic and anaerobic processes are handled by bacteria at the end of the food chain. From a trophic perspective, autotrophic bacteria transform matter, usually inorganic substances, in the presence of light, while heterotrophic bacteria utilize energy from complex organic compounds without light. During anaerobic processing, substances can be broken down into alcohol, methane, sulfides, and toxic byproducts, undesirable for the life of plants and fish in the aquarium. Therefore, it is essential to ensure an adequate supply of free oxygen in our aquariums. Bacteria mineralize matter, and this energy is transformed back into water and the root systems of plants, becoming a potential starting point for the cycle of substances once again.
Moja odborná špecializácia je environmentalistika, špecializáciu ekosozológia a fyziotaktika na Prírodovedeckej fakulte v Bratislave. Počas môjho štúdia som za zaoberal smerom ku geografii, konkrétne smerom ku Geografickým informačným systémom – GIS-om. Tento záujem neutíchol ani dnes. Musím povedať, že napriek svojmu terajšiemu aj minulému zamestnaniu som rád, že som vyštudoval environmentalistiku. Dostalo sa mi rozsiahlych vedomostí zo širokého spektra vedeckých disciplín. Okrem toho ešte dôležitejšie je, že to zodpovedá môjmu presvedčeniu. Environmentalistiku možno chápať ako autekologickú štúdiu, ako ekológiu zameranú na vzťahy organizmov, prostredia na človeka. V istých medziach je to veda, avšak myslím že poslanie environmentalistiky je v riešení situácií. Často sa v mnohých otázkach spája s medicínou, s ekonómiou a politikou. Ide o ochranu životného prostredia, ale len v súvislosti s človekom.
My professional specialization is environmental science, with a focus on ecozoology and physiotactics at the Faculty of Natural Sciences in Bratislava. During my studies, I also delved into the field of geography, specifically Geographic Information Systems (GIS). This interest persists to this day. I must say that, despite my current and past employment, I am glad that I chose to study environmental science. It has provided me with extensive knowledge across a wide range of scientific disciplines. Moreover, more importantly, it aligns with my conviction.
Environmental science can be understood as an autecological study, an ecology focused on the relationships between organisms and their environment, particularly concerning humans. In certain aspects, it is a science, but I believe the mission of environmental science lies in problem-solving. Often, it is intertwined with medicine, economics, and politics. It revolves around the protection of the environment, but always in the context of its impact on humanity.
Návrh na predmety vhodné pre environmentalistiku – praktický názor absolventa
Preferoval by som oveľa viac vzťahové vedomosti oproti encyklopedickým. Myslím, že dobrý ochranár nie je prioritne ten, kto vie, ale ten kto má vzťah. Ja by som sa tam snažil presadiť psychológiu, sociológiu, filozofiu, právo, ekonómiu. Na to by som kládol veľký dôraz. Tieto by mal študent ovládať a tie ostatné – prírodovedné by mali byť študentovým koníčkom. Z hľadiska praxe sa mi totiž zdá dôležitejšie či to vieš presadiť, nie či sa v tom vyznáš. Zbytoční sú ľudia, ktorí ovládajú veľké množstvo vedomostí, keď ich nevedia využiť. Úspešný adept si musí veriť.
Metodiky ako LANDEP a ÚSES, by som odporučil učiť len okrajovo. Vymysleli sa už stohy papierov, koncepcií, metodík, krajinnoekologické analýzy, syntézy, ale mnohokrát sú samoúčelné. Metodiky sú často také, že veľakrát by ju mal použiť človek, ktorý tomu nerozumie a ani nemá na to prostriedky, aby ju zodpovedne dodržiaval. Aj odborníkovi dajú slušne zabrať.
Suggestion for Subjects Suitable for Environmental Studies – Practical Opinion of a Graduate
I would prefer much more relational knowledge over encyclopedic knowledge. I believe that a good conservationist is not primarily someone who knows but someone who has a connection. I would try to emphasize psychology, sociology, philosophy, law, and economics in this field. I would place great emphasis on these. Students should master these, and the others – the natural sciences – should be the student’s hobby. From a practical perspective, it seems to me more important whether you can assert yourself, not whether you are knowledgeable. People who possess a vast amount of knowledge but cannot use it are unnecessary. A successful candidate must believe in themselves. I would recommend teaching methodologies like LANDEP and ÚSES only marginally. Many stacks of papers, concepts, methodologies, landscape-ecological analyses, syntheses have already been invented, but many times they are self-serving. Methodologies are often such that a person who does not understand them and does not have the means to responsibly adhere to them would use them. They can be quite challenging even for experts.
Separovaný zber a spotreba energie
Separujem zber asi od roku 1995. Keďže ma zaujíma koľko dokáže odpadu domácnosť vyprodukovať, a spotrebovať a za akých podmienok dochádza k poklesom a nárastu, zistené hodnoty môžem vyhodnotiť.
Zber papiera podľa rokov
V rokoch 1999/2000 som bol na vojenskej službe, preto zrejme došlo ku poklesu, nárast v roku 1998 vysvetľujem tým, že som bol denne doma. Nárast v rokoch 2001 až dodnes vysvetľujem veľkými nárastmi reklamných a informačných materiálov.
Separate Collection and Energy Consumption
I have been separating waste since around 1995. Since I am interested in how much waste a household can produce, and consume, and under what conditions there are fluctuations, I can evaluate the valuesI have found.
Collection of paper by years
In the years 1999/2000, I was on military service, so there was probably a decrease. The increase in 1998 is explained by the fact that I was at home every day. The increase from 2001 to the present is explained by significant increases in advertising and informational materials.
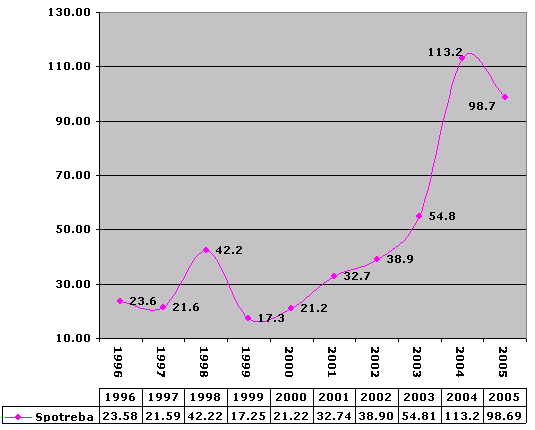
K 1.1.2006 som vyseparoval 472 kg papiera. /As of January 1, 2006, I separated 472 kg of paper.
Zber plastov
Na začiatku, keď sme začali separovať, veľmi dlho trvalo, kým si na to dostalo do vedomia, svoje zohrala aj ochota a zvyky. Potom, ako možno vidieť z roku 1998 sa situácia stabilizovala.
The separation of plastics was challenging at the beginning, and it took a while for awareness to build. Habits and willingness also played a role. As seen from the data in 1998, the situation stabilized over time.
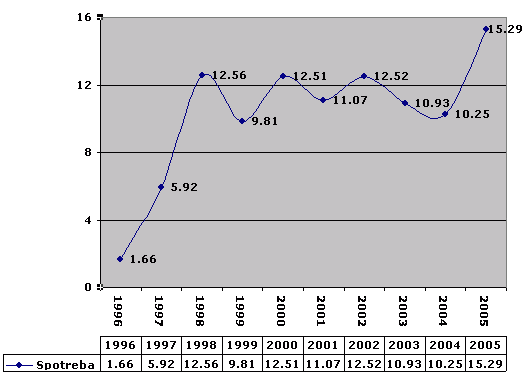
K 1.1.2006 som vyseparoval 105 kg plastov.
Kovy
Na príklade kovov je jasne vidieť veľké rozdiely, ktoré som naznačoval vyššie. V rokoch 1999 – 2000 som bol na ZVS. Konzervová strava sa jasne prejavila – okolo 7 kg na rok.
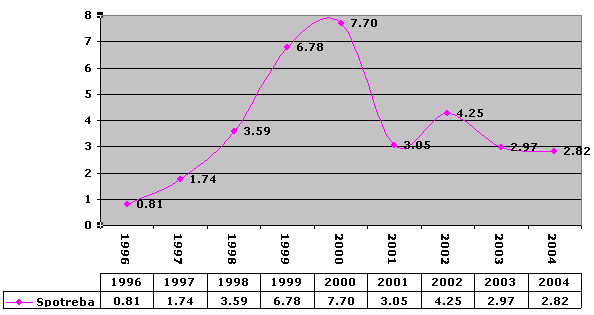
K 1.1.2006 som vyseparoval 46 kg kovov.
Spotreba energie
Evidované obdobie – od roku 1998
Voda
On January 1, 2006, I separated 46 kg of metals.
Energy Consumption
Recorded period – from 1998
Water
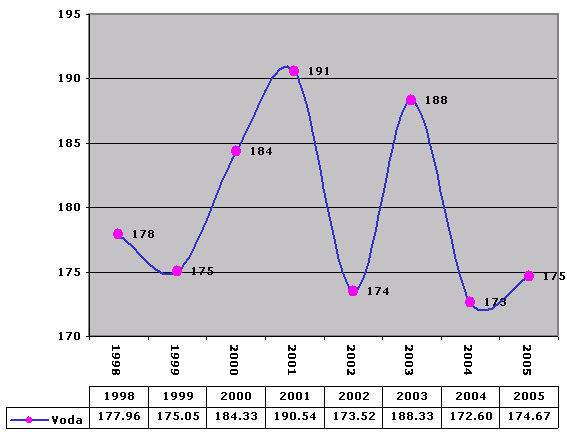
Spotreba je m3 /The consumption is in cubic meters.
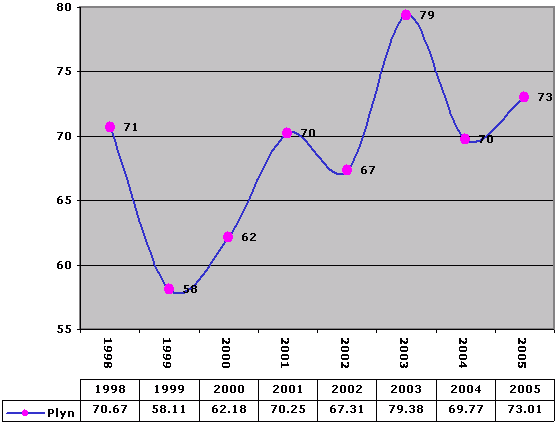
Spotreba je m3
Elektrina
Podľa jednotlivých rokov – v roku 2001 došlo ku výraznejšiemu nárastu – príčina najmä pomerne silné ohrievače – v rokoch 2002, 2003 došlo ku miernemu poklesu – vymenili sme starú chladničku za síce väčšiu a kombinovanú s mrazničkou, ale novú. V roku 2005 sme kúpili novú práčku, v roku 2006 nový sporák.
Electricity
By individual years – in 2001 there was a significant increase – the main cause was relatively powerful heaters – in 2002, 2003 there was a slight decrease – we replaced the old refrigerator with a larger one and combined with a freezer, but new. In 2005, we bought a new washing machine, and in 2006 a new stove.
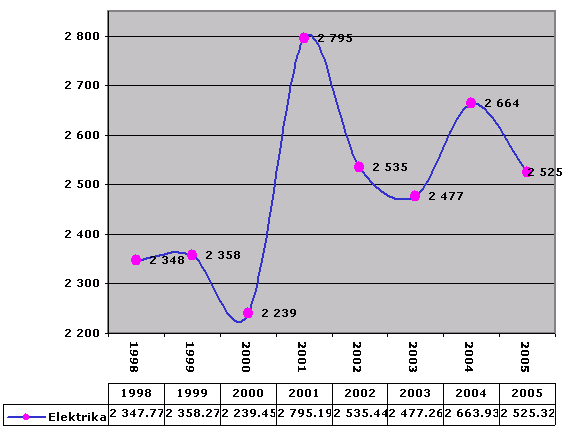
Spotreba je kWh.
Ekologická stabilita, ekologická rovnováha a spätná väzba predstavujú kľúčové pojmy v oblasti ekológie a environmentálnej vedy. Tieto koncepty sú úzko späté a vzájomne ovplyvňujú fungovanie ekosystémov. Ekologická stabilita sa vzťahuje na schopnosť ekosystému odolávať zmene a udržiavať svoje štruktúry a funkcie napriek vnútorným alebo vonkajším vplyvom. Stabilný ekosystém je schopný obnoviť sa po vonkajších narušeniach a udržiavať svoje rovnovážne podmienky. Ekologická rovnováha zdôrazňuje stabilitu v interakciách medzi živými a neživými časťami ekosystému. Táto rovnováha vzniká prostredníctvom komplexných vzťahov, kde organizmy ovplyvňujú svoje prostredie a súčasne sú ovplyvňované týmto prostredím. Spätná väzba je proces, pri ktorom výsledok alebo stav súvisí s predchádzajúcim stavom. V ekológii sa často používa na opis interakcií v rámci ekosystémov. Spätná väzba môže byť pozitívna (zosilňujúca) alebo negatívna (vyvažujúca). Napríklad, ak rastliny rastúce v ekosystéme absorbuje viac CO2, môže to viesť k pozitívnej spätnej väzbe v znižovaní celkového množstva skleníkových plynov v atmosfére. Úvaha o týchto konceptoch má kľúčový význam v súvislosti s globálnymi environmentálnymi výzvami. Súčasný stav našej planéty jasne ukazuje, že nevyvážené ľudské aktivity môžu narušiť ekologickú stabilitu a rovnováhu. Dôležité je uvedomiť si, že ekosystémy sú vzájomne prepojené a zmena v jednej oblasti môže mať významné dôsledky v iných častiach ekosystému. Z tohto dôvodu je potrebné konať s ohľadom na zachovanie a obnovu ekologickej stability, aby sme zabezpečili udržateľnú budúcnosť pre našu planétu.
Ecological stability, ecological balance, and feedback are key concepts in the field of ecology and environmental science. These concepts are closely interconnected and mutually influence the functioning of ecosystems. Ecological stability refers to the ability of an ecosystem to resist change and maintain its structures and functions despite internal or external influences. A stable ecosystem can recover from external disturbances and maintain its equilibrium conditions. Ecological balance emphasizes stability in interactions between living and non-living parts of the ecosystem. This balance arises through complex relationships where organisms influence their environment, and, at the same time, are influenced by it. Feedback is a process where the outcome or state is related to the previous state. In ecology, it is often used to describe interactions within ecosystems. Feedback can be positive (amplifying) or negative (balancing). For example, if plants in an ecosystem absorb more CO2, it can lead to positive feedback in reducing the overall amount of greenhouse gases in the atmosphere. Reflection on these concepts is crucial in the context of global environmental challenges. The current state of our planet clearly shows that unbalanced human activities can disrupt ecological stability and balance. It’s important to realize that ecosystems are interconnected, and a change in one area can have significant consequences in other parts of the ecosystem. Therefore, action is needed to preserve and restore ecological stability to ensure a sustainable future for our planet.
Abstrakt:
Tento článok sa zaoberá ekologickou stabilitou ako takou, a možnosťami jej zisťovania. Popisujem základné poznatky o ekologickej stabilite. Už v úvode uvádzam vzájomne pozitívny vzťah ekologickej stability a energie. Analyzujem možnosti jej zisťovania cestou produkčnej ekológie. Snažím sa popísať problémy s tým spojené. Pravdepodobne najzaujímavejším názorom celého článku je hypotéza, že závislosť ekologickej stability a energie nutnej na udržovanie momentálneho stavu je konštantná ! Osobitne sa venujem spätnej väzbe. Myslím, že spätná väzba má veľký vplyv na ekologickú rovnováhu. Schválne som použil slovo rovnováhu, pretože tak ako z článku vyplýva, mám veľké pochybnosti o jej vplyve na ekologickú stabilitu.
Abstract:
This article is dealing with ecological stability as the science, and possibilities of its detection. I describe the basic knowledge about ecological stability. I have mentioned other positive relationship between the ecological stability and the energy in introduction. I analyse the possibilities of its detection through productive (energy) ecology. I tried to describe troubles joined to it. I think the most interesting opinion of the whole article is a hypothesis that the dependence of ecological stability and energy which is necessary for sustaining of momentary state is constant ! I peculiarly devote to feedback. I think that feedback has a great influence on ecological balance. The article indicate that there is my deep doubt about the influence of feedback on ecological stability that is why I have just used the word balance.
Motto: Najnepochopiteľnejšou vecou na svete je fakt, že je to pochopiteľné (Albert Einstein)
Motto: The most incomprehensible thing in the world is that it is understandable. (Albert Einstein)
MÍCHAL ET VOLOŠČUK (1991) definujú ekologickú stabilitu ako schopnosť ekosystému vrátiť sa pôsobením vlastných vnútorných mechanizmov k dynamickej rovnováhe alebo ku svojmu “normálnemu” vývojovému smeru. Čím rýchlejšie sa ekosystém vracia a tým menšie odchýlky vykazuje, tým je stabilnejší. Rád by som zdôraznil, že ekologická stabilita je prevrátenou hodnotou vstupov, či už vo forme látok alebo energie, ktoré je nutné do ekosystému vkladať na to, aby sme ho udržali v momentálnom stave. Táto situácia nemá environmentálny aspekt. Neexistuje prepojenie na prirodzenosť ekosystému. Napr. lán pšenice vyžaduje každoročné mechanické narušovanie, hnojenie a kosenie, organický materiál (slama) musí byť odstraňovaný, alebo aspoň eliminovaný – inak by úroda v najbližších štyroch – piatich rokoch nebola takmer žiadna. Tok energie je prerušovaný.
Metodiky určovania koeficientu ekologickej stability v tvorbe ÚSES – Územného systému ekologickej stability sú do istej miery intuitívne. Je veľkým problémom zistiť mieru ekologickej stability presnejšie. Metodika je založená na znakoch, ktoré sa vyznačujú veľkým intervalom a veľkú úlohu zohráva subjektivita ľudského faktora. Projekty ÚSES vypracúvajú rôzni ľudia a s rôznym profesijným zameraním – od stavebných inžinierov, urbanistov, architektov cez krajinných plánovačov, biológov, ekológov až po environmentalistov. V tvorbe ÚSES nie je otázka presnej determinácie ekologickej stability taká dôležitá, okrem iného aj z dôvodu využívania princípu relatívnosti v tvorbe ÚSES. Prirovnal by som to k situácii lekára, ktorý sa snaží pomôcť pacientovi trpiacemu nevyliečiteľnou formou leukémie. Napriek tomu, že vie, že pacientovi veľa času neostáva, hľadá oporu – “kostru”, najhodnotnejšie časti, ktoré pacienta udržia v stave najvyššej možnej rovnováhy, prípadne pocitu spokojnosti. Toto prirovnanie trochu kríva, totiž príroda nikdy nebude v stave pacienta, avšak na pochopenie relatívnosti využitia poznania ekologickej stability pre účelovú prácu akou ÚSES je vhodný.
MÍCHAL ET VOLOŠČUK (1991) defines ecological stability as the ability of an ecosystem to return, through its internal mechanisms, to dynamic equilibrium or its “normal” developmental direction. The faster an ecosystem returns and the smaller the deviations it exhibits, the more stable it is. I would like to emphasize that ecological stability is the inverted value of inputs, whether in the form of substances or energy, that need to be invested in the ecosystem to maintain it in its current state. This situation does not have an environmental aspect. There is no connection to the naturalness of the ecosystem. For example, a wheat field requires annual mechanical disruption, fertilization, and mowing; organic material (straw) must be removed or at least eliminated — otherwise, the crop in the next four to five years would be almost nonexistent. The flow of energy is disrupted.
The methodologies for determining the coefficient of ecological stability in the creation of ÚSES – Územný systém ekologickej stability are, to some extent, intuitive. It is a significant challenge to ascertain the degree of ecological stability more precisely. The methodology is based on features characterized by a wide range, and subjectivity plays a significant role. ÚSES projects are carried out by various individuals with different professional backgrounds – from civil engineers, urban planners, and architects to landscape planners, biologists, ecologists, and environmentalists. In the creation of ÚSES, the exact determination of ecological stability is not as crucial, partly due to the use of the principle of relativity in its formation. I would liken it to the situation of a doctor trying to help a patient suffering from an incurable form of leukemia. Despite knowing that the patient has limited time, the doctor seeks support, a “framework,” the most valuable parts that will keep the patient in the highest possible state of equilibrium or a sense of satisfaction. This analogy is somewhat skewed because nature will never be in a patient’s state, but it helps understand the relativity of using knowledge of ecological stability for the purposeful work that ÚSES represents.
Čo je to ekologická stabilita a aké sú možnosti jej zisťovania a akéhosi “ocenenia” pre laikov ? To je zložitá otázka, na ktorú sa snažím dať nejaké podnety. Môžem použiť dokonca medicínu, či ekonómiu na to, aby som dokázal pozitívny vzťah ekologickej stability k cieľom uvedených odborov. Ak poviem ekonómovi, že toto územie má takú a takú cenu z hľadiska ekologickej stability, a že na tom svojou správne orientovanou aktivitou (napr. pasením na lúke v Strážovských vrchoch) ušetrí ročne istú sumu peňazí, je vhodné, aby sme vedeli určiť ekologickú stabilitu. Je všeobecne známy pozitívny vzťah rovnováhy v prírode k zdraviu obyvateľov. Metodiky zisťovania koeficientu ekologickej stability s ktorými som sa najčastejšie stretol sú založené na získavaní poznatkov o aktuálnom stave vegetácie na jednotlivých plochách. Vegetácia je reprezentatívnym ukazovateľom, ktorý dostatočne presne odráža stav ekosystému nielen v momentálnom čase, ale je reakciou na podmienky okolia počas pomerne dlhého časového obdobia. Tieto metodiky majú jeden veľmi exaktný aspekt – je ním pôvodnosť. Napr. drvivá väčšina lúk ako takmer nepôvodný prvok v našich podmienkach nedosahujú najvyšší význam pre ekologickú stabilitu – hodnotu 5, ako takmer všetky ostatné typy ekosystémov . To je dané nevyhnutnosťou vstupov energie pre ekosystém lúk, ktoré musia mať zabezpečený manažment na to aby pretrvali vo svojej forme. Keď som uvažoval o presnejšom postupe pre zisťovanie stupňa ekologickej stability usúdil som, že pravdepodobne najexaktnejšia metóda merania stupňa ekologickej stability je cesta využívajúca metódy produkčnej ekológie. Lenže to má dva hlavné okruhy problémov.
Prvý problém je v tom, že jednotlivé ekosystémy majú rôznu výšku produkcie, a rôznu výšku potrebných vstupov. Druhý problém spočíva v nasledujúcom. Téza hovorí: ekologická stabilita je nepriamo úmerná energii nutnej na udržovanie momentálneho stavu (MÍCHAL ET VOLOŠČUK, 1991). Ako sa dá zistiť koľko nevyhnutnej energie treba. Je možné pomerne presne vyčísliť energiu, ktorá je do ekosystému vkladaná človekom, ale ako možno odhadnúť aké množstvo je nutné na udržanie rovnakého stavu? Okrem toho ekosystém sa každoročne mení, niektoré druhy gradujú, každý rok je iný prísun zrážok, napriek tomu sa stabilita nemusí zmeniť. Na ekologicky nestabilných plochách by tieto problémy narastali.
Je známe, že existuje závislosť medzi ekologickou stabilitou, energiou nutnou na udržovanie momentálneho stavu a energiou dodávanou do ekosystému prirodzene. Myslím si dokonca, že táto závislosť je konštantná pre všetky typy ekosystémov. V akom vzťahu je energia dodávaná do ekosystému umelo ku energii dodávanej prirodzene vzhľadom na jeho stabilitu ? Dnes vieme len tom, že v nepriamo úmernom. Veď napokon je to logické, však ? Akým spôsobom je možné zistiť tento vzťah ? Ja sa nazdávam, že cesta je schodná práve prostredníctvom produkčnej ekológie. Veľkým problémom však je pravdepodobne zistiť presné množstvo energie, ktoré je nutné pre prirodzený vývoj najmä v neprirodzených ekosystémoch. Ak by sa zistili energie a vypočítal by sa ich vzájomný podiel, mohol by sa porovnať s metodikou zisťovania ekologickej stability používanou v tvorbe ÚSES a potom už by nemal byť väčší problém matematickým aparátom s veľkou presnosťou odhadnúť vzťah ekologickej stability a energie.
What is ecological stability and what are the possibilities of determining it and some kind of “reward” for laypeople? This is a complex question that I am trying to provide some insights into. I can even use medicine or economics to demonstrate a positive relationship between ecological stability and the goals of these fields. If I tell an economist that this area has a certain value in terms of ecological stability and that, through properly oriented activities (such as grazing in the meadows of the Strážovské vrchy mountains), a certain amount of money can be saved annually, it is appropriate to be able to determine ecological stability. The generally known positive relationship between balance in nature and the health of the population supports this idea. Methods for determining the coefficient of ecological stability that I have encountered most frequently are based on acquiring knowledge about the current state of vegetation on individual areas. Vegetation is a representative indicator that accurately reflects the state of the ecosystem not only at the present time but is a response to environmental conditions over a relatively long period. These methods have a very precise aspect — originality. For example, the vast majority of meadows, as an almost non-original element in our conditions, do not achieve the highest importance for ecological stability — a value of 5, like almost all other types of ecosystems. This is due to the necessity of energy inputs for meadow ecosystems, which must have management in place to survive in their form. When I considered a more precise approach to determine the degree of ecological stability, I concluded that probably the most accurate method for measuring the degree of ecological stability is a path utilizing methods of production ecology. However, this has two main circles of problems.
The first problem is that individual ecosystems have varying levels of production and different levels of required inputs. The second problem is as follows. The thesis states: ecological stability is indirectly proportional to the energy necessary to maintain the current state (MÍCHAL ET VOLOŠČUK, 1991). How can one determine how much necessary energy is required? It is possible to fairly accurately quantify the energy being input into the ecosystem by humans, but how can one estimate the amount needed to maintain the same state? Additionally, the ecosystem changes every year, some species graduate, there is a different annual precipitation influx, yet stability may not change. On ecologically unstable areas, these problems would escalate.
It is known that there is a dependence between ecological stability, the energy needed to maintain the current state, and the energy naturally supplied to the ecosystem. I even believe that this dependence is constant for all types of ecosystems. In what relationship is the energy artificially supplied to the ecosystem to the naturally supplied energy concerning its stability? Today, we only know that it is inversely proportional. After all, it is logical, isn’t it? How can we determine this relationship? I think the way is through production ecology. However, a significant problem is probably determining the exact amount of energy needed for natural development, especially in unnatural ecosystems. If the energies were determined, and their mutual ratio calculated, it could be compared with the methodology used to determine ecological stability in the creation of ÚSES. Then, there should be no major problem to estimate the relationship between ecological stability and energy with great precision using mathematical tools.
Spätná väzba veľmi výrazným spôsobom vplýva na ekologickú stabilitu. Je to vzájomné, nenáhodné pôsobenie medzi prvkami (prípadne subsystémami) toho istého systému, pri ktorom dochádza k zosilňujúcemu – pozitívnemu, alebo zoslabujúcemu – negatívnemu pôsobeniu veličiny B, ktorá bola priamo, alebo nepriamo zmenená veličinou A, na túto veličinu A. Spätná väzba je najdôležitejším autoregulačným mechanizmom všetkých systémov bez výnimky (MÍCHAL ET VOLOŠČUK, 1991, p. 28), a preto ma vzťah k ekologickej stabilite. Dôkaz takejto spätnej väzby prináša užitočné informácie o chovaní systému (okrem iného o jeho stabilite) i v prípade, že mechanizmus tohto pôsobenia pre nás ostáva neznámy (MÍCHAL ET VOLOŠČUK, 1991, p. 28). Pri pozitívnej spätnej väzbe pôsobí každý z dvoch premenných prvkov v interakcii zhodným smerom, takže sa navzájom zosilňujú v pozitívnom, či negatívnom zmysle. Napr., čím viac členov populácie A je v plodnom veku, tým viac potomstva B sa môže narodiť. Potom, čím viac potomstva B dorastá do plodného veku, tým rýchlejšie narastá populácia. Dochádza teda k posilňovaniu obidvoch premenných prvkov interakcia v čase a výsledkom je známa exponenciálna rastová krivka tvaru “J” (MÍCHAL ET VOLOŠČUK, 1991, p. 28 – 29). Pozitívna spätná väzba posilňuje odchýlky a nerovnovážne stavy a spravidla slúži dynamickému rastu systému. Ten však časom naráža na určitú hranicu – limity svojho vonkajšieho prostredia. Kvalitatívne zmeny vlastností prvkov interakcie sa potom môžu prejaviť principiálne dvoma spôsobmi (MÍCHAL ET VOLOŠČUK, 1991, p. 28 – 29):
Patologické javy vyvolané stresom sú v spätnoväzbovej schéme faktorom “naviac” k schéme pozitívnej väzby. S rastúcim počtom plodnej časti populácie narastajú nielen počty potomstva, ale progresívne rastie nedostatok potravy a priestoru a ďalších zdrojov nevyhnutných pre existenciu ďalších členov populácie, prípadne rastie znečisťovanie prostredia samotnou populáciou. Tieto rastúce negatívne vplyvy v prostredí populácie prinášajú jej jedincom takú záťaž, že sa to prejaví patologickými zmenami chovania, ktoré môžu viesť i v prírodných podmienkach k prudkým poklesom populačnej hustoty smerom k minimu (MÍCHAL a VOLOŠČUK, 1991). Z toho vyplýva, že pozitívna spätná väzba, najmä vo vrcholových častiach krivky spôsobuje zníženie stupňa ekologickej stability. Pýtam sa, či môže mať spätná väzba ako prírodný a prirodzený proces autoregulácie ekosystémov negatívny vplyv na ekologickú stabilitu ako schopnosť ekosystému ? Napr. v takom ekosystéme ako je tajga je registrované pravidelné výrazné kolísanie početnosti vtákov a stavovcov. Napriek tomu stabilita tohto ekosystému je vysoká, jeho schopnosť vysporiadať sa s vnútornými i vonkajšími faktormi prostredia je obdivuhodná. Charakteristika pomerov v tajge môže viesť k názoru, že daný ekosystém je nevyvážený, rozkolísaný, jednoducho že nie je v stave ekologickej rovnováhy. Avšak tento ekosystém pokiaľ nie priamo ohrozovaný človekom sa vyznačuje vysokým stupňom ekologickej stability. Mechanizmus pretrvávajúceho zabezpečovania rovnováhy je pravdepodobne spätná väzba. MÍCHAL ET VOLOŠČUK (1991) hovoria o tom, že ak fluktuácia zložiek ekosystému nevedie k dlhodobej deštrukcii jeho štruktúr, môže byť kolísanie početnosti určitej populácie výrazom ochranných aktivít riadených spätnými väzbami. Spätná väzba je prostriedkom regulácie, môže spôsobiť ekologickú rovnováhu, ale pochybujem o tom, že negatívne vplýva na ekologickú stabilitu.
Feedback significantly affects ecological stability. It is a mutual, non-random interaction between elements (or subsystems) of the same system in which there is an amplifying – positive, or attenuating – negative influence of variable B, which was directly or indirectly changed by variable A, on this variable A. Feedback is the most important self-regulating mechanism of all systems without exception (MÍCHAL ET VOLOŠČUK, 1991, p. 28), and therefore, it is related to ecological stability. Evidence of such feedback provides useful information about the behavior of the system (including its stability) even if the mechanism of this action remains unknown to us (MÍCHAL ET VOLOŠČUK, 1991, p. 28). In positive feedback, each of the two variable elements in the interaction acts in the same direction, so they mutually reinforce each other positively or negatively. For example, the more members of population A are in the fertile age, the more offspring B can be born. Then, the more offspring B grows into the fertile age, the faster the population grows. Thus, both variable elements reinforce each other in time, resulting in the well-known exponential growth curve in the shape of “J” (MÍCHAL ET VOLOŠČUK, 1991, p. 28 – 29). Positive feedback strengthens deviations and unbalanced states and generally serves the dynamic growth of the system. However, over time, it encounters a certain limit – the limits of its external environment. Qualitative changes in the properties of interaction elements can then manifest themselves in two fundamentally different ways (MÍCHAL ET VOLOŠČUK, 1991, p. 28 – 29):
Patological phenomena induced by stress are an additional factor in the positive feedback loop. With the increasing number of the fertile part of the population, not only the numbers of offspring increase, but there is a progressively growing shortage of food, space, and other resources necessary for the existence of other members of the population, or pollution of the environment increases due to the population itself. These growing negative impacts in the population’s environment impose such a burden on its individuals that it manifests as pathological changes in behavior, which can lead, even in natural conditions, to sharp declines in population density towards the minimum (MÍCHAL and VOLOŠČUK, 1991). Therefore, positive feedback, especially in the upper parts of the curve, causes a reduction in the degree of ecological stability. I wonder if feedback as a natural and inherent process of ecosystem self-regulation can have a negative impact on the ability of the ecosystem, such as the taiga, to maintain stability? For example, in the taiga, there is regularly significant fluctuations in the abundance of birds and mammals. Despite this, the stability of this ecosystem is high, and its ability to cope with internal and external environmental factors is admirable. The characteristics of relationships in the taiga may lead to the opinion that the ecosystem is unbalanced, fluctuating, simply not in a state of ecological balance. However, as long as it is not directly threatened by humans, this ecosystem exhibits a high degree of ecological stability. The mechanism of maintaining balance is probably feedback. MÍCHAL and VOLOŠČUK (1991) state that if the fluctuation of ecosystem components does not lead to the long-term destruction of its structures, the fluctuation in the abundance of a particular population may be an expression of protective activities regulated by feedback. Feedback is a means of regulation; it can cause ecological balance, but I doubt that it negatively affects ecological stability.
Literatúra
MÍCHAL, I., ET VOLOŠČUK, I., 1991: Rozhovory o ekológii a ochrane prírody, ENVIRO, Martin.